

HOME  論文/ハイライト 研究ハイライト 研究者インタビュー 柘植謙爾講師
論文/ハイライト 研究ハイライト 研究者インタビュー 柘植謙爾講師
08.10.21
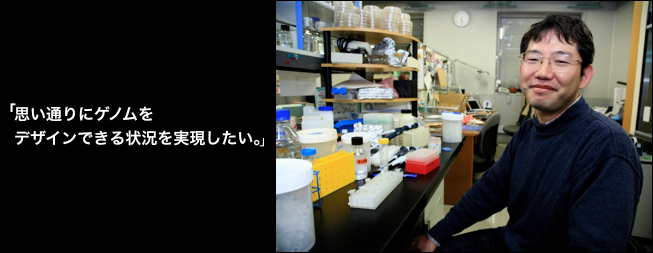
柘植謙爾講師
(08.10.21)

─現在はどのような研究テーマに取り組まれているのでしょうか。
一番大きな目的は『ゲノムをデザインする』ことです。ゲノムデザインというと、生物をつくる、というニュアンスに近いのかもしれませんが、それよりも限定的に、DNAを操って生物に必要な完全なゲノムを作るにはどうしたらいいのか、ということを目標にしています。
その中でも、トップダウン方式とボトムアップ方式という2つの方式があります。トップダウンは板谷さんがやっていらっしゃる研究で、ある生物の中に違う生物のゲノムを完全に入れこんでから、徐々に外来ゲノムの方を発現させるためにはどうすれば良いか、という方式です。
僕がメインに考えているのはボトムアップという方式で、遺伝子の単位からゲノムをデザインしていくという発想です。ボトムアップ方式で非常に重要な実験手法として遺伝子集積があります。遺伝子の断片を自分で操れないとゲノムデザインは難しいので、効率良く遺伝子を集積する方法を発明してゲノムをデザインしていきたいな、というわけです。
─その方法とは?
枯草菌プラスミドの形質転換系を利用して遺伝子集積をする方法で、名前をOGAB法(Ordered Gene Assembly in Bacillus subtilis法)と言います。この方法は、僕が知る限り、という前提がつくのですが、少なくともゲノムをデザインするという観点においては最も優れた方法だと考えています。
一番成績が良いものでは、13個のDNA断片を混ぜてライゲーションという連結操作を1回するだけで、自分が指定した順番通りに全断片を連結することができました。しかも、OGAB法は薬剤マーカーでプラスミドを持っている枯草菌の形質転換体をとってくるのですが、8割以上が13個の断片を正しい順序や向きに連結していたという状況でした。
ゲノム構築において遺伝子の並べ方は重要か?
─13個のDNA断片の向きや順序にはどのような工夫をされているのでしょうか?
13個の断片中に遺伝子が10個あったのですが、10個の遺伝子の連結順序というのは10の階乗通りあります。10の階乗ってどのくらいになると思います?すぐには出てこないと思いますが、362万8800通りという数です。オペロンという一つの転写単位を作成する時にそういう数が発生するということです。遺伝子がコードしている向きはすべて左から右へと一定の方向なのですが、その向きの中での並び順というのが約300万通り存在するわけですね。ちなみにそのコードの向きを考慮せずに並べ替えをしたとすれば、単純な意味でいくつになるのですかね・・・さらに2の10乗倍かかってくるのですぐには計算できませんが、もっと莫大な数字になります。遺伝子の向きを一定にして考えただけでも300万という数字になるわけです。これらの遺伝子を連結したものが、果たして同じような性質を示すのか、というところに非常に興味を持っています。
たとえば大腸菌には4千数百の遺伝子がありますが、その4千数百の遺伝子をただやみくもにつなげて一本のDNAにしたら生物として成り立つのかといえば、そんなことはないと考えています。順番というのが非常に重要な概念として成立しているのではないか、それを解き明かしたい。ということで、手はじめに、その遺伝子をどういう順番で連結すればよいか、順番に意味があるのかについて研究しようとしています。
─うまくいったプラスミドの遺伝子発現はどうだったのでしょうか?
今までに論文にした成果では、カロテノイドという、皆さんが食品でとるような色素性物質の5つの遺伝子を使いました。5 つの遺伝子には5の階乗通りの並び順が存在しますが、代謝(経路)で遺伝子がはたらく順序の早いものからABCDEという風に並べて1つ目の並び順にしました。次はEが先頭に来てEABCD、その次はDEABCという形で、先頭を順に移動させた5種類の並び順を選びました。これにプロモーターをつなげてオペロンにし、 大腸菌で発現可能なプラスミドとして枯草菌でクローニングしてから大腸菌に導入してカロテノイドの生産量を調べました。すると、遺伝子を連結する順番によって生産量が変わるという現象が見いだされ、代謝の順序に並べたときに一番生産量が高いという結果が得られました。(T. Nishizaki et al., 2007, Applied and Environmental Microbiology)。
こういう現象を何が規定しているのかについては非常に興味がありました。そこで、各遺伝子のmRNAのその時点で残って いる存在量、つまり(生産されたもの)-(分解されたもの)の量をRT-PCR法で測定してみました。その結果、遺伝子の連結順にかかわらず、mRNAの 量が最も多いのはプロモーターに近いところに連結された遺伝子でした。一方、mRNA量が少ないのはプロモーターから遠い遺伝子で、連結順にしたがって単調に減少していました。つまり、mRNAの存在量は一回下がってから増加したり中央が高くなるのではなく、プロモーターの位置から単純に減少していくこと が分かりました。
野生型のオペロンでもそうなるのではないかと漠然と言っている論文はありますが、遺伝子の連結順序によっては、遺伝子の構成に変化がなくても活性(発現量)が変わってしまう現象があることになります。すなわち、遺伝子の単位から生物を作り上げようとした時には、遺伝子を連結する順番は非常に重要ではないか、ということです。
─10個の遺伝子をつなげたときも、代謝の順につなげた物が機能としては一番良かったのでしょうか?
10個の遺伝子は解糖系の遺伝子なのですが、代謝の順に連結したものをまず作りました。次に、これらの遺伝子は大腸菌由 来のものなのですが、中東さんがとられた野生型の大腸菌における各解糖系遺伝子のmRNA量のデータをみると、10個の遺伝子の中に非常に多く発現してい る遺伝子と発現していない遺伝子があることが分かったので、そのデータに基づいて並べたオペロンを作りました。代謝順のものとmRNA量の順番のものをよく見るとほぼ正反対に近いような並び方をしていて、代謝経路の先頭の遺伝子は発現量が比較的少なくて、経路の後半にくる遺伝子は発現量が多いことが分かり ました。そこで、代謝経路にしたがったものを中央で切って逆転させたものも作ってみました。
この3つのオペロンの性質を調べるのは難しく、解糖系がない生物に入れれば一番良いのですが大腸菌の解糖系遺伝子には必須遺伝子が多いので、作ったオペロンの機能を完全に評価することはなかなかできませんでした。そこで、IABにあるKeioコレクションという各大腸菌の1遺伝子を欠損した株に作ったオペロンを持つプラスミドを入れて増殖を調べました。どの欠損株でも、あるプラスミドを入れて普通に生息したらそのプラスミドが100%働いているのだろうという推測ができます。
その結果、最初につくった代謝経路順に並べたプラスミドではある程度増殖は回復したものの、野生型ではmRNA量が多い 代謝経路後半に位置する遺伝子の欠損株はあまり相補できませんでした。プラスミドを入れたことで本来はコロニーを得られない培地で生えるようにこそなりま したが、増殖が非常に遅いという状況でした。一方、代謝経路前半の転写量をあまり必要としない遺伝子の欠損株は完全に回復していました。
それを改善しなければ遺伝子の連結順序を解決したことにはならないので、野生型大腸菌のmRNA発現量を基準にして並べ たものについて同じ実験をしたところ、一遺伝子欠損株は全て完全に相補したのです。ほぼ野生型と同じように生育できるようになりました。なので、ひょっとすると、遺伝子が必要としているmRNA量の多い順番に遺伝子を連結することに意味があるのではないかと考えているところです。
─エネルギー生産に近い遺伝子がより多く発現している、ということなのでしょうか?
それは非常に難しいのですが、たとえば先ほど説明したカロテノイドではそういう傾向はなく、代謝経路の順番にしたがっていたものが比較的成績が良かったので、個々のケースによるのではないかと見ています。カロテノイドの遺伝子に関しては野生型のmRNA量を調べていないの で、というか、そもそもカロテノイドを作る遺伝子が大腸菌の遺伝子ではないので調べようがないのですが、大腸菌の解糖系遺伝子については調べることができ ます。野生型のオペロンについては調べることができませんが、ともかく必要としているmRNA量の順番にしたがって並べる手法は代謝経路がわかっていなくても利用できるという点において非常に有用だと考えています。
─OGAB法では一度にいくつまで遺伝子をつなげることができるのでしょうか?
集積する遺伝子の断片が突出を持つように調整します。この突出は、DNA二本鎖の3'末端が飛び出るような形で、かつ、出っぱっている部分が3塩基になるように設計をしています。
3塩基の配列の相補な関係にあるものはリガーゼをいれると連結できますが、逆の言い方をすれば、相補な関係のものが一カ 所以外には出現しないように突出の形状を連結部位の数だけ用意してあげれば、いくつもの断片を一度に連結できます。この3塩基の突出は4種類の塩基からなっているので、4の3乗で64通りあるのですが、右の断片と左の断片というかたちで相補なものも64個の中にふくまれているため、実際には32カ所の連結ができることになります。ただし、経験上なのですが、GやCをふくまないAとTだけの突出は水素結合が非常に弱いためになかなかうまくいかないなど、突出の形によってはライゲーションが難しい配列もあります。そういうことも考慮すると、断片の数としてはせいぜい20個位が限度かもしれません。しかしながら、3塩基の突出を5'末端突出で作ることも不可能ではないので、同じ配列でも突出の形状の違いによってコントロールできれば、原理上は無制限に連結でき るのではないかと思います。
もう少し具体的な話をしますと、この方法が今まで他の菌、たとえば大腸菌などでできなかった理由があります。枯草菌は DNAを自発的に取り込む性質がありますが、大腸菌では電気をかけたり、塩化カルシウムの溶液につけて膜構造を変化させたりすることでDNAを無理矢理入れています。
形質転換が得意な枯草菌を使ったからできたこと。
枯草菌がDNAを取り込むときは細胞表層にDNAを認識するタンパク質が出現して、2本鎖のDNAを捕まえて1本鎖になるよう切断した後、DNA の片方の鎖だけを菌の中に取り込みます。もうひとつの鎖は細胞表層で分解されているらしく、菌体中に入るのは1本鎖のDNAです。それを考えると、プラスミドは環状の2本鎖DNAなので、そんなものが枯草菌の細胞表層で捕まえられて切断されてしまったら菌の中でプラスミドがもとの形に戻るということはあまり期待できません。実際、大腸菌のようなプラスミドの形質転換は非常に難しかったですね。

ところが、枯草菌でプラスミドの形質転換ができないというわけではなくて、1回取り込んだ1本鎖のDNAをまた元の環状プラスミドに再生するような配列があれば簡単に形質転換ができます。その配列のことを我々はタンデムリピートと言っているのですが、あるプラスミドの ユニットが全く同じ方向に複数連結した直線状のDNAがあると、これはどこかで細胞表層から菌体の中に取り込まれていくのですが、そのときに1本鎖で入っていったプラスミドが1個以上入ると、切断された部分をリカバーするような情報をもったDNAというのが入っていきます。詳しいメカニズムはわかりません が、切断箇所をリカバーするような情報を利用してその部分を直すことを枯草菌はしていて、それを利用してプラスミドを菌体の中で再生してしまう、ということができるみたいです。一見不思議なのですが、こういう形でプラスミドの形質転換ができることは昔から分かっていました。
要するに・・・大腸菌で複数の断片からなるプラスミドを作ることは極めて困難なのですが、枯草菌の形質転換ではDNAが切断されることが大前提なので、環状DNAを作る必要がありません。線状になっていればよくて、かつ繰り返しがあればどこで切断されるか分からな いランダムな切断なんですけれども、それをリカバーするためには1個分以上入ればよいと。その部分を利用すると、複数の断片からなるプラスミドを簡単に再生できるわけです。こうして、遺伝子集積が可能になることを発見してこれを使おうとしたのですが、技術的な部分でひとつだけ問題があります。大体おわかりいただけると思うのですが、DNA断片のモル数がすべて同じ比率である必要があります。たとえば、あるベクターだけがあるものに対して2倍多いとすると、 両端がベクターで止まってしまってその先の連結が続きません。それでも多分集積はできると思いますが、DNA断片の濃度がバラバラだときれいなタンデムリピートはできません。このDNAの濃度を合わせるところが技術的には難しいです。断片の数が多くなるほど高い効率を出すのが難しいのですが、DNAの濃度を正確に量る測定方法である程度解決できるということが分かってきました。
完全なプラスミドを切断して断片比を揃える・・・逆転の発想で成功。
DNAの濃度をコントロールするときに、最初は電気泳動で写真をとって濃度を調べていたのですが、その後蛍光物質による正確な定量を行うように なってはじめて、先ほど説明したような13断片もの接続が可能になってきました。一回集積して完成したプラスミドは突出を作るために制限酵素を利用してつくっているので、人工的な制限酵素サイトが残っています。よって、同じ制限酵素で切断するともとのDNA断片に戻るのですが、それを再度集積に用いるわけです。最初につくったのは人工的に調整した断片なので幾らかモル比にズレがあるのですが、2回目には一度完成したプラスミドを切断して得られた断片なのでモル比がきれいに1になります。それをもう一度連結し形質転換すると、自分が一生懸命濃度比をあわせた物よりも100倍とか1000倍のオーダーで形質転換頻度が高くなることがわかっています。そこからも、濃度を綺麗にあわせることがこの方法において非常に重要である、ということが分かります。
─非常にスマートな方法ですね。
僕もそう思います(笑) 非常に簡単な原理ですが、これを実際にやってみたところがすごい。そして思った通り非常にうまくいったということですね。この方法の特徴をもう少しだけ宣伝させて頂きますと、これは最近論文にしたものなのですが、0.6kbしかない小さな遺伝子断片と、 1.0kbくらいの遺伝子断片、そして38kbある大きな遺伝子断片を用意しました。大きな断片と小さな断片の長さの比は64倍くらいと、非常に大きな違いがあります。これらの断片と10数kbあるプラスミドの断片を集積したところ、大きな遺伝子に対して小さな遺伝子が同じ向きかどうかなど、さまざまな向きの物をつくることに成功しました(K. Tsuge et al., 2007, Journal of Biotechnology)。この方法では遺伝子の長さに関係なくプラスミドを構築できるというメリットがあります。大腸菌のプラスミドは環状にする必 要がありますが、おしりと頭が空間的に出会う可能性が極めて低くなるため、長くなればなるほど環状化が難しくなります。枯草菌で連結をする場合は、長さ自体は関係なくいくらでも伸びます。この違いによって、集積の効率が極端に違ってきます。これは基本的には大腸菌では無理な方法で、枯草菌を使ってはじめて できる点で我々が優位に立っていると言えます。
─強く影響を受けた人や、出来事はありますか。
大学院は東京工業大学の試験科学研究所というところで、現在の化学農薬(化学的に合成された農薬)に比べて環境に優しいと考えられる微生物農薬というものの研究開発を行っていました。具体的にはコンポスト、要するに堆肥です。ある堆肥にいる微生物が抗生物質を出して病原菌の作用を抑えている、という事実が当時分かってきたところで、僕はその抗生物質の遺伝子の研究で博士号をとりました。その抗生物質の遺伝子はサイズが40kbもある極めて長いものですが、非常に大きな遺伝子を取り扱う技術が当時はまだなく、大きな遺伝子を自由に加工する技術が必要だと感じていました。 その後、今の三菱化学生命科学研究所で、大きな遺伝子を取り扱うための技術を習得したいということで板谷さんのところに入ったという経緯があります。なので、枯草菌をずっと扱っていましたし、大きなDNAもずっと扱っていました。あまり研究の断絶はなくここまで来ているというような状況です。
─一番最初に手がけた研究は何ですか。
学部時代は埼玉大学の伏見譲先生という方の研究室にいました。伏見譲先生はものすごく有名な先生で、レーザーと蛍光物質を利用してシーケンシングを自動化する機械の基本的な原理を考案した人です。残念ながら特許は取れなくて、のちのち大きな損失であったことが明らかになるわけですが・・・
そこでは進化分子工学をやっていて、人工的に進化が起こせるんじゃないかっていう、そういうものを検証するための実験系をやっていました。大腸菌のlacZ遺伝子をできるだけ大量に発現させるようプロモーターに変異を加え、lacZの発現量をモニターするための蛍光物質を取り込ませます。細胞の中でその蛍光物質をいっぱい出させてセルソーターという機械にかけると蛍光強度をモニターすることができるので、lacZ発現量の高いものを細胞一個のレベルで分離できます。そうしてとってきた優良な株のプロモーターに対してさらに変異をかけて、同じ操作を繰り返していくと、人工的な進化が検証できるわけです。蛍光物質をどのように大腸菌に取り込ませるか、いったん取り込ませた蛍光物質が酵素の活性によって蛍光を持つようになった後、大腸菌から漏れないようにするにはどうしたら良いかという研究をしていました。そういうところからも人工的な進化に関する興味があって、今でも関心として持っています。
─欠かせない日課はありますか。
最近、鶴岡に来てからはよく釣りをするようになりました。その行為自体は暇なところもありますが、自然の中に入ってゆっくりと今までの研究を振り返るよい時間になっているのではないかと思います。鶴岡という自然環境にいるからこそ、可能だということですね。非常に優れたアイディアが思いついたというわけではありませんが。
─「封神演義」の太公望みたいな感じですね(笑)
そうですね、常に大物を狙っていきたいということで(笑) そういう研究方針でやっています。
─それでは最後に、将来の展望をお聞かせください。
ゲノムシーケンスというのが非常に行われてきて、遺伝子の種類や配列はやたらとたくさん手に入れることができました。しかしながら、そのままでは単にテキストの情報にしかすぎなくて、それを有効活用した例はほとんどありません。自分たちはそれらの情報を実物に変換するための橋渡しができたらいいなぁ、と思います。
一つの遺伝子を連結していくことで、モノをたくさん作るようなシステムを構築したいと思っています。たとえばバイオエタノール等の燃料の問題や、あるいは二酸化炭素排出の問題がありますが、そういうものをクリアできるような基礎技術開発の一端を担えたらいいなと。ゆくゆくはそれをさらに発展させて、完全に自分の思いどおりにゲノムをデザインできる状況が実現できたらいいなと考えています。
─本日はどうもありがとうございました。
(2007年11月10日 インタビューア:小川雪乃 編集:西野泰子 写真:増田豪)